





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
结核分枝杆菌调节miRNAs及circRNAs抗细胞自噬机制的生物分析
[张益源1 , 罗嘉婧2 , 伊正君2 , 付玉荣1  ]
]
]
|
|


目的 探索结核分枝杆菌抵抗人或小鼠巨噬细胞自噬机制的关键miRNAs及circRNAs,为寻找新型结核诊断标志物及治疗靶点提供思路。方法 筛选出结核分枝杆菌诱导细胞差异表达的自噬相关基因;制作蛋白交互图并文献挖掘筛选关键自噬基因;多数据库综合预测相应的miRNA,制作自噬基因mRNA-miRNAs交联图并筛选关键miRNA;制作miRNAs-circRNAs相互作用图并筛选关键circRNA。结果 筛选出18种结核感染后差异表达的自噬相关基因,并在此基础上预测16种自噬关键基因;筛选出4种通过靶向自噬mRNA参与自噬调控的关键miRNA分子(miR-17-5p, miR-20a-5p, miR-30a-5p, miR-33-5p);针对4种miRNA筛选出6种与其密切相关的circRNA分子,并预测hsa-circRNA2908和hsa-circRNA6439是影响结核抗自噬关键circRNA分子。结论 miR-miR-17-5p, miR-20a-5p, miR-30a-5p, miR-33-5p及其上游共同调控分子hsa-circRNA2908和hsa-circRNA6439可能是潜在的诊断标志物和疾病治疗靶点。
We explored the pivotal miRNAs and circRNAs which can affect the Mycobacterium tuberculosis (MTB) resistance to the human or mice macrophage autophagy, to provide the new way to find the novel tuberculosis diagnosis biomarker and therapeutic target meanwhile. We screened and filtrated the differentiated expressed autophagy related genes regulated by MTB in infected macrophage and selected the pivotal autophagy genes from the protein-protein interactions diagram and literature mining. We also integrated forecasting the relevant miRNA with that through multiple predicting databases, create the autophagy gene mRNA-miRNAs interaction diagram and selected the pivotal miRNA, verifying them by literature or experiments. Then we created miRNAs-circRNAs interactions diagram and selected the pivotal circRNA likewise. Of 18 autophagy related genes with differentiated expression in MTB infected cells were filtrated, and 16 crucial autophagy genes were predicted in the basis. Moreover four pivotal miRNAs (miR-17-5p, miR-20a-5p, miR-30a-5p, miR-33-5p) were selected which regulate autophagy by targeting mRNA of autophagy genes. In addition, six relevant potential circRNAs were selected by virtue of analysis of the connection with that four mRNAs, and speculating that has-circRNA2908 and has-circRNA6439 would be the pivotal circRNA to impair MTB resistance to cell autophagy. MiR-17-5p, miR-20a-5p, miR-30a-5p, miR-33-5p and their common upstream regulating molecules, has-circRNA2908 and has-circRNA6439 may be potential diagnosis biomarker and therapeutic target of disease.
结核分枝杆菌(Mycobacterium Tuberculosis, MTB)是结核病的病原体, 因具有超强的抵抗力而在机体内难以被消灭, 不断刺激机体发生炎性反应并逐渐形成结核肉芽肿从而发展成慢性疾病, 其较强的生存力和抗巨噬细胞自噬、逃避免疫杀伤等机制有密切关系。自噬是指吞噬了胞内成分的囊泡和溶酶体融合形成自噬溶酶体, 降解其所包含的内容物的过程[1]; 自噬可以清除自身蛋白质、细胞器, 还可以清除胞内感染的微生物, 与炎症相关, 是十分重要的细胞生物学现象[2]。自噬相关基因是指能表达参与细胞内自噬过程的相关蛋白的一系列基因, 其表达程度受microRNA(miRNA)的调节。miRNA是一种长度约18~22 nt的内源性非编码单链RNA分子, 通过靶向基因的3'UTR区域, 降解靶基因或抑制其翻译, 在转录后水平调节基因的表达[3, 4]; miRNA的表达水平又受其上游分子Circular RNA(circRNA)、Long non-coding RNA(lncRNA)的调节。circRNA不具有5'末端帽子和3'末端poly(A)尾巴, 是以共价键形成环形结构的非编码RNA分子[5]。circRNA起miRNA海绵作用, 其内含miRNA应答元件并可与之结合, 阻止miRNA抑制靶基因, 上调相应基因表达[6]。 近年来研究发现多种miRNA在肺结核患者体内显著升高或降低, 其通过靶向巨噬细胞内自噬相关基因表达从而抑制自噬, 增强MTB的抗自噬能力和生存力; 而实验抑制相关miRNA的表达能够显著增强细胞自噬清除MTB的能力, 因此探索潜在的相关miRNA及其调控分子在疾病的诊断和治疗中具有巨大意义。本文依据文献报道及实验分析预测了与结核抗自噬相关的自噬基因, miRNAs, circRNAs, 并阐述其相互作用通路, 为进一步研究结核分枝杆菌通过调控miRNAs、circRNAs抗细胞自噬奠定理论基础。
利用GEO数据库 (https://www.ncbi.nlm.nih.gov/geo/) 检索结核感染相关实验数据, 筛选4个实验样本进行分析比较[7, 8, 9, 10]。选择样本的标准为结核分枝杆菌肺部感染人或小鼠, 实验细胞为巨噬细胞, 实验感染时间长。对每个样本的基因表达情况进行分析, 制作Volcano Plot图(火山图)。从目标结果中筛选自噬相关基因, 多个样本结果选取交集, 初步得到差异基因[11]; 用GeneMANIA工具[11] (http://genemania.org)、String网站[12](https://string-db.org) (形成差异基因间蛋白质交互作用(PPI)网络模型, 观察这些自噬基因的PPI作用方式及联系程度, 用Cytoscape软件 (www.cytoscape.org)[13]对网络模型数据加工处理, 制作基因间相互作用网络交联图, 将交联图中联系较为密切的自噬蛋白群筛选出来, 再合并一些通过文献挖掘得到的自噬基因, 组成最终差异的自噬基因群; 并用GCBI网站 (https://www.gcbi.com.cn/gclib/html/index) 制作多基因雷达关联网络图将基因群联系起来。
选择基因雷达群中存在广泛联系的差异自噬基因, 采用miRWalk 2.0 (http://http://zmf.umm.uni-heidelberg.de/apps/zmf/mirwalk2/index.html)[14]软件进行Gene-miRNA分析。为预测靶向自噬基因的miRNA准确度, 选择TargetScan、miRanda、miRDB、miRWalk、RNAhybrid共5个数据库预测的miRNA结果交集做为预测结果。筛选条件设置为P< 0.05, 最小种子序列长度为7 mer, 靶基因结合区域为3'UTR。得到的数据结果用Cytoscape软件进行处理及图形绘制, 制作最终的Gene-miRNAs交联网络图。差异自噬基因-miRNA网络分析选出与自噬基因交联多的miRNA, 文献挖掘验证miRNA的表达差异, TargetScan (http://www.targetscan.org/vert_71/)[15]、miRanda (http://34.236.212.39/microrna/home.do)[16]检验miRNA与靶基因保守型。
用starBase v2.0 (http://starbase.sysu.edu.cn/index.php)[17]的“ miRNA-circRNA” 工具预测筛选出来的miRNA的上游circRNA分子, 用Cytoscape软件制作miRNAs-circRNAs交联图。利用交联图取各miRNA预测结果交集, 得到相关circRNA分子。结合文献挖掘, 选出表达发生显著差异的circRNAs作为参与自噬调控的关键circRNA分子。
相关数据采用SPSS 24.0软件进行分析处理, 统计方法采用t检验, P< 0.05为差异具有统计学意义。
样本的Volcano Plot图分析结果见图1, 筛选条件为P< 0.05, 表达下调2倍以上, 目标结果用橙色显示。 结果中挑选出的自噬相关基因, 以KEGG数据库 (http://www.genome.jp/kegg/pathway.html) Autophagy-animal通路中已有的基因为标准。GeneMANIA工具、String网站分析发现这些基因表达蛋白间还存在非常复杂的相互作用, 参考KEGG、Reactome (https://reactome.org)[18]的自噬通路发现在MTB感染后表达显著变化的自噬基因功能主要集中在自噬囊泡的形成过程, 表现出很高的特异性。从Volcano Plot图表达差异显著的基因中挑选出稳定保守的自噬相关基因, 用String网站分析得到这些基因表达蛋白交联作用图, 并用Cytoscape细节处理后结果见图2:更加直观的展示了参与MTB调节自噬的相关基因及基因间的联系。差异表达的自噬相关基因文献挖掘结果见表1:这些自噬基因在MTB感染后发生显著上调或下调, 并且都已实验证实受miRNA调节[19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33]。从图2中筛选出10个自噬相关基因(BECN1, ATG3, ATG4b, ATG5, ATG7, ATG12, ATG14, ATG16L1, UVRAG, WIPI1), 合并6个经文献挖掘的自噬相关基因(ATG10, ULK1, DRAM1, VMP1, LAMP3, MAP1LC3A), 最终筛选出的基因群的雷达网络交联图见图3。
 | 图1 MTB感染的细胞内差异表达基因火山图 过滤阈值:差异倍数FC> 2, |log2(FC)|> 1; P< 0.05, -log10 (P)> 1.30; 中间蓝色的分割线表示FC为1; 分割线左边表示下调的基因, 右边表示上调的基因; 橙色区表示表达下调倍数> 2的基因Fig.1 The volcano plot of the differentiated expression of genes in MTB-infected cells |
 | 图2 MTB感染细胞内差异自噬基因的相互作用网络 基因节点的大小和颜色深浅表示其交联作用的复杂程度Fig.2 The interaction network of differentiated autophagy related genes in MTB-infected cells |
 | 图3 自噬基因间相互作用雷达网络 浅蓝色的点表示自噬蛋白, 黄色的点“ GABARAP” 表示其和自噬蛋白的交联作用最复杂Fig.3 The autophagy genes in gene radar of interaction network |
基因-miRNAs网络分析图结果见图4; 将表中与基因交联数较多(≥ 3)的miRNA挑选出来, 基本情况见表2; 部分miRNA在MTB感染后的表达上调或下调[34, 35], 部分miRNA与自噬基因的作用关系在其他非结核领域实验中已经验证[36]。文献挖掘出已实验证实的结核感染细胞后差异表达的自噬基因-miRNA相互作用关系, 详细结果见表1。
| 表1 MTB感染细胞内受miRNA调节的自噬基因文献挖掘结果 Tab.1 The literature outcome of autophagy genes regulated by miRNA in MTB-infected cells |
| 表2 筛选出的miRNAs和自噬相关基因间的相互作用 Tab.2 The interaction information between the filtered miRNAs and autophagy related genes |
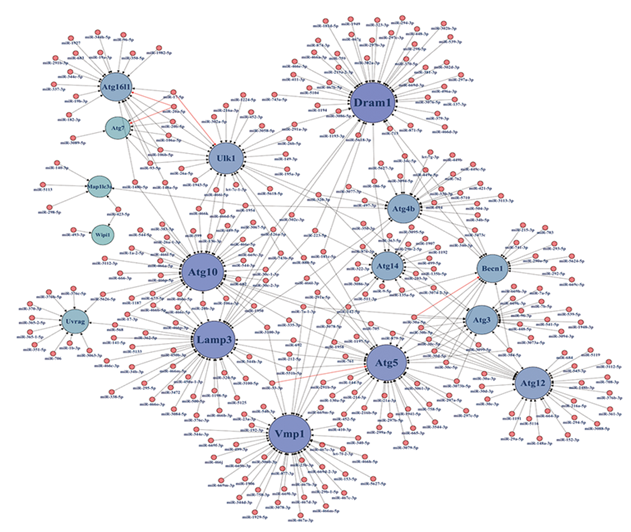 | 图4 差异自噬基因-miRNAs相互作用网络 基因节点的大小和颜色深浅表示其与其他miRNA交联作用的复杂程度, 已经被实验证实的miRNA和基因的相互作用用红线表示Fig.4 The interaction network between differentiated autophagy genes and miRNAs |
经验证的miR-17-5p, miR-20a-5p, miR-30a-5p, miR-33-5p共4种可信度高的miRNA, 预测对应的circRNAs, 筛选阈值为最高可信度(very high stringency≥ 5), 4种miRNAs的交联结果见图5(a)。交联后共得到6个同时靶向4种关键miRNA的circRNA(图5(a)最上处的6个紫色节点代表该6种circRNA), 分别为SHOC2_hsa-circRNA1108, UBE3C_hsa-circRNA7333, RPA2_hsa-circRNA 8391, GIGYF1_hsa-circRNA15246, AFF4_hsa-circRNA14411, CLTC_hsa-circRNA3463, 见图5(b)。此外交联图中的SLC30A7_hsa-circRNA414, UBFD1_hsa-circRNA2908, CAMLG_hsa-circRNA6439已经被证实在结核感染相关实验中表达显著差异, 具体结果见表3[37, 38, 39, 40]。将筛选阈值调整到中等时(medium stringency≥ 2), 发现circRNA686、circRNA1937、circRNA7851、circRNA12825靶向性强且表达差异显著, 但circRNA686不利于MTB抗自噬; 此外hsa-circRNA6439还靶向miR-30a-5p, 其抗自噬功能尤为显著。综上, 将整个通路参与自噬调控的重要分子整理归纳, MTB抗细胞自噬的整个调控过程“ MTB-Gene-miRNA/circRNA-Gene” 示意图见图6。
 | 图5 miRNAs-circRNAs间的相互作用网络^(a)miRNAs与circRNAs交联网络整体布局; (b)框架内的紫色节点表示差异表达的关键circRNAsFig.5 The interaction network between miRNAs and circRNAs |
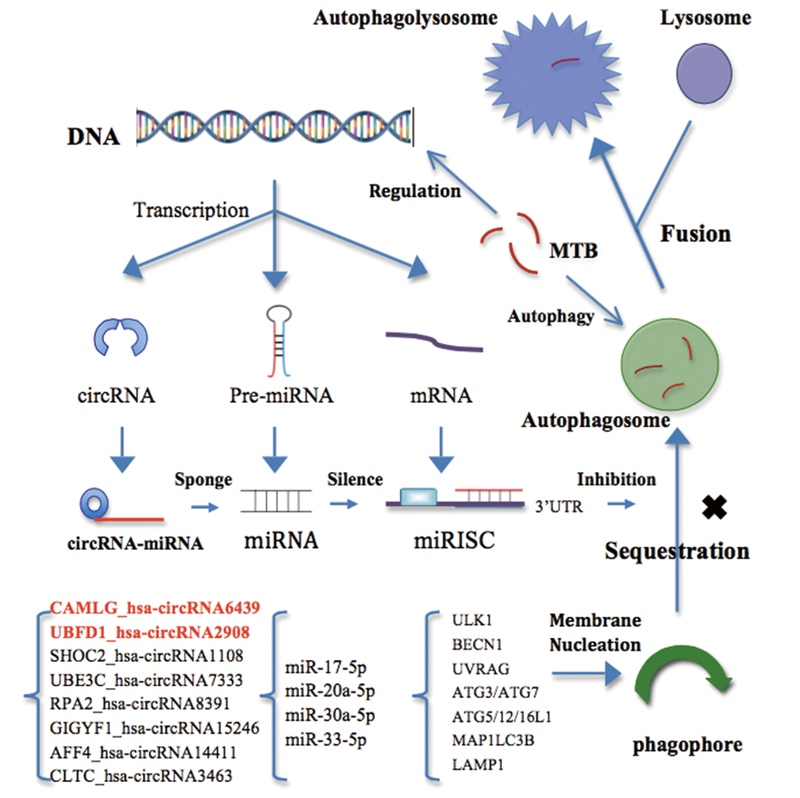 | 图6 MTB抗巨噬细胞自噬机制图 红色字体表示的circRNAs是关键RNAFig.6 The schematic diagram about MTB resistance to the autophagy in macrophage |
结核病是由结核分枝杆菌感染引起的慢性传染病, 以肺部感染最为常见。近几年来结核病越来越难以治疗, 这和其本身出现耐药性及顽强的抵抗力密切相关。结核分枝杆菌抗细胞自噬是增强其在细胞内生存力的重要机制, 其能阻止细胞自噬体的形成或逃避自噬溶酶体吞噬从而一直存在于细胞内免于杀伤[41]。通过大量的生物信息学分析及实验验证, 我们发现感染结核分枝杆菌后细胞有大量自噬相关基因表达下调, 其中有明显差异的基因共18个, 见图2。经过大量文献挖掘, 从中选出一些可靠的自噬基因并增加一些其他实验验证的自噬基因, 组成共16个自噬关键基因, 见图3(雷达中蓝色节点代表的基因)。通过差异自噬基因-miRNAs交联网络图我们发现了miR-17-5p, miR-20a-5p, miR-30a-5p, miR-33-5p共4种可信度较高的关键miRNA, 且这些miRNA和自噬高度相关且有实验报道[20, 21, 24, 25, 26]。通过miRNA-circRNAs交联网络图发现了6种潜在的circRNA及2种重要的circRNA(见图5b), 作为miRNA上游分子它们同时靶向上述4种miRNA, 抑制miRNA的基因沉默作用, 在自噬作用中很可能起重要作用。
图1显示MTB感染巨噬细胞后样本中大量基因表达下调, 这和MTB某些成分对巨噬细胞的基因表达调控有关。实际上, MTB被巨噬细胞吞噬进入细胞质内, 通过自身特异蛋白或分泌蛋白(如ESAT-6等)作用于宿主基因组, 调节相关转录因子表达等进而调整其他基因表达, 直接调节自噬基因或先调节miRNA进而间接控制自噬相关基因的表达[42, 43]。如表1所示, 近几年研究发现miRNA在MTB抗细胞自噬方面有重要作用, 人和小鼠细胞实验结果也基本一致。如图6所示, 结核杆菌可能通过上调上述4种miRNA, miRNA又形成RNA沉默复合体(miRISC)靶向自噬基因的mRNA, 导致mRNA降解从而翻译自噬蛋白[44]。细胞缺少自噬蛋白便无法合成吞噬泡(Phagophore), 成膜过程发生障碍从而自噬体(Autophagosome)减少, 溶酶体(lysosome)无法和自噬体融合, 自噬溶酶体(Autophagolysosome)合成下降, MTB逃避吞噬最终生存下来[45]; 相反自噬恢复就会增强MTB的清除[46]。而circRNA能够抑制miRNA, 发挥miRNA海绵的作用, 从而阻止miRNA抑制自噬的作用, 有些差异表达的circRNA还是潜在的诊断标志物[37, 38]。在图5(a)中靶向miR-30a-5p的SLC30A7_hsa-circRNA414, 分别靶向miR-20a-5p及miR-17-5p的UBFD1_hsa-circRNA2908和CAMLG_hsa-circRNA6439三个分子已被相关实验证实, 在活动肺结核血中分别上调2倍以上, 5倍以上和10倍以上; 其中UBFD1_hsa-circRNA2908, CAMLG_hsa-circRNA6439尤为敏感, hsa-circRNA6439还能靶向miR-30a-5p而增强自噬, 它们很有可能是结核领域调控自噬的关键circRNA成员及相关诊断标志物[37, 39]。另6种多交联的circRNA在自噬调节方面可能也发挥重要作用, 但还需要进一步实验验证。目前有关MTB和circRNA的调控作用关系仍不是很清楚, circRNA在巨噬细胞感染MTB后的表达变化和其在细胞内的具体作用有待更进一步的研究, 本文为研究其抗自噬相关作用提供一定思路。参与自噬调控的上述4种miRNA都已实验证实在MTB感染的细胞内表达含量发生显著差异, 且在人体血清样本也已证明含量显著变化, 这为利用miRNA作为潜在的新型无创诊断生物标志物提供一定依据[20, 21, 24, 25, 26]。此外, 将关键circRNAs和miRNAs作为药物作用的治疗靶点, 通过上调或下调相关分子从而增强细胞自噬功能, 巨噬细胞清除MTB能力大大将会增强, 这为寻找药物作用靶点提供新思路和重要依据。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|