




{kind=link}
{kind=link}
{kind=link}
{kind=link}
登革热病毒(DENV)包膜蛋白特异性人源抗体筛选与鉴定
[高博1, 2 , 郑晖2 , 李枢2 , 谢联辉3  ]
]
]
|
|


目的 建立自外周血快速筛选识别登革热病毒衣壳的人源抗体的方法,从病毒感染者外周血记忆B细胞中筛选出特异性抗体并进行性质分析。方法 通过流式细胞术分选得到外周血中登革热病毒包膜蛋白特异性的单个记忆B细胞,结合高效单细胞PCR技术获得抗体基因序列,对重组表达的单克隆抗体进行结合活性及中和活性检测。结果 成功筛选到识别DENV-1包膜蛋白的6株人源单克隆抗体,其中4株抗体均具有很强的结合活性以及中和活性。结论 成功的通过流式细胞术对DENV特异性记忆B细胞进行分选,并结合单细胞PCR技术,实现了对DENV特异性人源单克隆抗体的高效筛选。通过该平台有希望筛选得到DENV的广谱中和抗体,用于登革热的预防和治疗。
We amied to establish a method for rapid screening of human antibodies against dengue virus(DENV) capsids from peripheral blood, and to screen specific antibodies from peripheral blood memory B cells of virus-infected patients for qualitative analysis. By flow cytometry, DENV envelope protein-specific memory B cells in peripheral blood were single-cell sorted. The gene sequence of antibody was amplified using efficient single-cell PCR. We determined the binding activity and neutralization potency of recombinant full-length antibodies. Six human monoclonal antibodies recognizing DENV-1 envelope protein were successfully screened, and four of them had strong binding activity and neutralizing activity. DENV-specific memory B cells were successfully sorted by flow cytometry, and DENV-specific human monoclonal antibodies were efficiently obtained using single-cell PCR. The results revealed that it is hopeful to obtain DENV-specific antibody with broadly neutralizing activity for the prevention and treatment of dengue fever.
登革病毒(DENV)属于黄病毒属的单股正链RNA病毒, 以埃及伊蚊和白纹伊蚊为传播媒介。DENV基因组全长约10~11 kb, 只有1个开放读码框, 共编码3个结构蛋白: 衣壳蛋白(C)、膜蛋白(M)、包膜蛋白(E)和7个非结构蛋白( non-structuralprotein, NS1~NS5)[1, 2]。DENV包膜(E)蛋白由3个结构域组成:E蛋白结构域1(ED1)是中心结构域, ED2是二聚化结构域并含有保守的融合环, ED3是推定的受体结合结构域[3]。 E蛋白占据病毒表面的大部分并且是体液免疫应答靶向的主要抗原[1, 4]。使用小鼠单克隆抗体进行的表位研究已经在E蛋白的表面上鉴定了至少12个不同的表位, 其中 ED3上的表位能够激发起有效的中和抗体[4]。其中4种血清型(DENV1-4)的感染可导致患者出现轻度登革热发热、严重的登革热出血热和登革热休克综合征等症状, 有些则成为无症状感染者[5, 6]。DENV感染主要在热带和亚热带地区流行, 在美洲、东地中海、东南亚和西太平洋地区报告的发病率最高。大多数登革热流行都是由单一血清型病毒引起的, 偶有报道由两种或更多血清型引起流行[7]。 DENV的4种血清型也可以在同一地区引起登革热流行病, 已有报道我国广东省暴发过4种DENV血清型的流行, 其中以DENV-1流行为主[8, 9, 10]。在过去的几十年, 全球约有4亿人每年感染登革热病毒, 其中约有1亿人出现不同程度的症状[11]。在医疗保健系统发达的国家中, 登革热的死亡率非常低。然而, 登革热的经济负担很高, 最近估计登革热治疗的全球成本为每年80亿至90亿美元[12]四价减毒活疫苗(Dengvaxia)已经成在许多国家获得许可[13]。然而, 登革热血清阴性的接种人群, 在之后的登革热感染时会出现明显的病情加重, 事实上, 这种疫苗现在已在菲律宾停止使用。因此, 需要针对该病毒的有效治疗方法。
抗体治疗正在成为一种针对病毒感染的潜在治疗方法。例如帕利珠单抗, 其是人源化的小鼠单克隆抗体, 基于儿童的临床试验, 发现其显着降低由于RSV感染而住院的风险[14]。目前, 多种技术手段都可应用于人源单克隆抗体筛选, 其中记忆B细胞分离后培养及单细胞PCR技术应用最为广泛[15, 16, 17]。此外, 记忆B细胞永生化技术也发展的十分迅速, 已经成功应用到肿瘤特异性抗体的筛选中。近期的研究中, 通过应用登革热疫苗接种者或病毒天然感染者的浆细胞进行单细胞PCR, 成功筛选得到多种人源登革热病毒特异性单克隆抗体, 具有应用于临床治疗的潜力[18, 19]。在本研究中, 我们使用流式细胞分选技术(FACS)筛选得到登革热感染者外周血单核细胞中的抗原特异性记忆B细胞, 通过单细胞PCR技术获得抗体基因序列, 在此基础上纯化表达获得了6株特异性人源抗体。对获得的抗体性质进行了初步的鉴定, 为开发高效的登革热检测方法及治疗药物提供了技术支持。
1.1.1 病例 该感染者在2014年广州市登革热暴发流行中被确诊, 于确诊后40 d采集外周血样本10 mL于抗凝采血管中。通过梯度密度离心分离外周单核细胞(PBMC), 储存于液氮中备用。4种型别重组E蛋白为本实验室生产。
1.1.2 主要试剂 常用质粒pMD18-T、pTT-5为实验室保存, 其中pTT-5载体已构建抗体轻重链恒定区, 用于抗体表达; 常用菌株如DH5α 、Top10购自Tiangen公司; 细胞株BHK-21、293Expi购自ATCC; DMEM培养基、胎牛血清、PBS磷酸盐缓冲盐溶液、青霉素、链霉素均购于美国Hyclone公司。样品的核酸和质粒提取试剂购自QIAGEN公司; 羊抗人IgGHRP(Fab specific)购自PIERCE公司; Live/dead-Aqua、CD3-PeCy7、CD20-FITC、CD27-PE、IgG-BV421及链亲和素偶联的APC等染色用荧光抗体购自BD公司, DENV-1 E蛋白生物素化试剂购自Thermo公司。抗体纯化介质购自GE公司; DNA、Protein Marker购自Thermo公司。
1.1.3 Nested-PCR引物 本研究所涉及的引物均由Invitrogen公司合成, 具体引物设计方案参照文献[16]。
1.2.1 单细胞分选 采集2014年广州市登革热暴发流行中被确诊的感染者外周血10 mL, 采用常规Ficoll-Paque密度梯度离心, 获得外周血单核细胞(PBMC)。采用Live/dead、CD3、CD20、CD27、IgG、IgM和DENV-1 E-biotin这6个指标对抗原特异的记忆B细胞进行特异性标记, 经BD AriaⅢ 分选型流式细胞仪进行数据采集和单细胞分选, 用96孔U底细胞板进行细胞收集。
1.2.2 逆转录PCR 在96孔PCR板中进行逆转录操作。配置14 μ L逆转录体系, 各组分加入量为:150 ng随机引物(pd(N)6, GE Healthcare)、0.5 μ L dNTP(Invitrogen)、0.5%Igepal CA-630 (Sigma)、4 U RNAsin (Promega)、6 U Prime RNAseInhibitor (Eppendorf) 和50 U Superscript○R III reverse transcriptase (Invitrogen), 42 ℃, 10 min; 25 ℃, 10 min; 50 ℃, 60 min; 94 ℃, 5 min。逆转录结束后, 将PCR板保存于-80 ℃冰箱。
1.2.3 Nested-PCR 通过Nested-PCR 对IgH, Igλ 和Igκ 可变区基因进行扩增, 扩增体系为40 μ L, 各组分够成为:cDNA 3.5 mL、20 nmol/L各引物、300 nMdNTP (Invitrogen) 和 1.2 UHotStar○R Taq DNA polymerase (Qiagen)。Nested-PCR反应条件为:94 ℃, 30 s; 58 ℃(IgH/Igκ ) 或者 60 ℃(Igλ ), 30 s; 72 ℃, 55 s (第一轮PCR) 或者45 s (第二轮PCR); 循环数50。应用2%琼脂糖核酸电泳, 对Nested-PCR产物进行鉴定及回收。
1.2.4 PCR产物纯化和克隆鉴定 Nested-PCR产物片段大小为500 bp, 使用TIANGEN回收试剂盒纯对鉴定为阳性的PCR产物进行回收, 然后克隆至pMD18-T载体。每个PCR产物挑取5个菌斑进行菌液PCR验证, 提取阳性克隆质粒进行基因测序。
1.2.5 抗体基因序列分析 将测序结果用MEGA 7读取, 并将测序结果正确的序列在IMGT数据库中进行重链以及轻链的家系及突变情况等分析。
1.2.6 抗体的表达及纯化 对纯化得到的重链及轻链可变区分别构建酶连位点, 将来自每个单细胞的抗体可变基因的扩增cDNA克隆到含有人IgG1或Igκ 恒定区的表达载体中, 构建不同轻重链表达克隆, 重组表达IgG1亚型抗体[20]。使用LipofectamineTM 2000转染试剂盒(11668-019, Invitrogen), 用等量的编码重链和轻链的质粒转染293Expi细胞。培养5 d后, 收集含有抗体的细胞上清, 并使用Protein A介质(17-1279-03, GE Healthcare)进行纯化。
1.2.7 抗体结合活性测定 通过不同型别重组E蛋白的酶联免疫吸附试验(ELISA), 对单克隆抗体与不同血清型DENV的结合活性进行检测。包被E蛋白(DENV-1:228-11688, DENV-2:228-11689, DENV-3:228-11690, DENV-4:228-11691; RayBiotech)200 ng每孔于ELISA板, 4 ℃过夜保存。然后每孔中加入200 μ L 5%脱脂奶粉, 37 ℃下孵育2 h进行封闭。呼吸道合胞病毒(RSV)特异性人源单克隆抗体作为阴性对照抗体, 其与待检测单克隆抗体以初始浓度为30 μ g/mL进行4倍倍比梯度稀释, 然后加入100 μ L各浓度, 并在37 ℃下孵育1 h。PBS-Tween洗涤3次, 加入羊anti-人IgG抗体(1∶ 5000, v/v), 37 ℃下孵育1 h, PBS-Tween洗涤3次。加入显色底物, 37 ℃孵育15 min, 终止显色并读取吸光度参数。根据读值, 计算其抗体与抗原的结合活性。
1.2.8 蚀斑减少中和试验(PRNT)测定抗体中和活性 将BHK-21细胞(5× 105细胞/mL, 2 mL培养基)接种于6孔板中, 37 ℃ 5%CO2条件下培养过夜。然后加入单克隆抗体-病毒混合液于BHK-21细胞单层中, 37 ℃下孵育1 h。弃去培养上清液, 加入800 μ L 1.2%营养甲基纤维素, 37 ℃下培养4~5 d。空斑形成后, 用0.1%结晶紫染色30 min, 观察斑块形成。汇总数据分析不同抗体中和活性。
为了获得天然的轻重链抗体配对, 本研究应用流式细胞分选系统, 实现单个B细胞高效分选。本研究中分析的样本来源于2014年广东省登革热感染者, 应用CD3、CD20、CD27、IgG、IgM及DENV-1 E蛋白等指标实现对PBMC中B细胞亚群的具体分析。记忆B细胞作为体液免疫应答的重要组成, 成熟记忆细胞表面的BCR具有很强的抗原结合能力且长期存在于外周血中, 因此本研究也选择记忆B细胞作为获得DENV抗原特异性单克隆抗体基因的来源。
分析结果表明, 外周血中成熟记忆B细胞(Aqua-/CD3-/CD20+/CD27+/IgG+)中DENV E蛋白特异性成熟记忆B细胞占比为0.32%, 如图1。从10 mL外周血的单核细胞中共分选得到110个特异性单B细胞, 用于后续的单细胞PCR。
 | 图1 DENV特异IgG+记忆B细胞群分析Fig.1 Analysis of DENV-specific IgG+memory B cells |
抗体轻重链可变区为抗原特异性识别区域, 通过获取该区域基因序列信息实现对抗体的序列进行分析及表达鉴定。已经广泛使用的单细胞PCR技术能够对IgH、Igλ 和Igκ 的可变区基因进行高效扩增。研究中单细胞PCR产物经琼脂糖凝胶电泳鉴定后, 显示在分选得到的110个DENV特异性B细胞中成功调取重链的有55个, 成功率为50%, 轻链的有70个(48个Kappa型, 22个lambda型), 成功率为63.6%。共得到24株抗体轻重链基因配对, 配对成功率为21.8%。采用Gibson装配方法将配对抗体轻重链可变区序列构建到含有人源IgG1或Igκ 恒定区的表达载体中。经过酶切鉴定及序列比对, 最终成功构建24株抗体的表达克隆。配对抗体轻重链表达质粒等量转染到293Expi真核表达细胞, 悬浮培养5 d后收集细胞培养上清。经过酶联免疫吸附试验(ELISA)鉴定, 其中6株为DENV特异性单克隆抗体。通过Protein A介质对IgG1抗体进行纯化, 纯化产物经离心浓缩后进行SDS-PAGE电泳鉴定。结果如图2, 各抗体的轻重链均可很好的表达, 重链分子量大小为50 kD左右, 轻链分子量大小为25 kD左右, 与正常大小相符合。
 | 图2 SDS-PAGE凝胶电泳分析抗体轻重链表达Fig.2 Antibodies were characterized by SDS-PAGE |
通过重组E蛋白的酶联免疫吸附试验(ELISA), 对单克隆抗体与不同血清型DENV E 蛋白的结合活性进行了检测, 对EC50进行统计分析。结果如图3, 这6株单克隆抗体具有DENV结合活性, 其均为DENV-1 E蛋白特异性单克隆抗体, 在高浓度下与DENV-2 E 蛋白具有很弱的结合。在6株单克隆抗体中, 4株单抗N-1、N-2、N-3和N-6具有与DENC-1 E蛋白相近的结合活性, EC50分别为0.006 9 μ g/mL、0.006 8 μ g/mL、0.005 4 μ g/mL和0.006 5 μ g/mL。此外, N-4与N-5的结合活性较差, N-5的EC50为2.06 μ g/mL, 这应该与其识别表位的差异性或者抗体成熟度不够有关。
通过蚀斑减少中和试验(PRNT)测定各抗体中和活性, 结果如图4。因为筛选得到的单克隆抗体为DENV-1特异性单克隆抗体, 所以其只对DENV-1病毒具有中和活性, 对其他亚型均无中和能力。普遍认为中和活性与结合活性之间具有一定的相关性, 本研究的中和结果也说明了这一观点。6株抗体中强结合活性的N-1、N-2、N-3和N-6均表现出很强的中和DENV-1病毒的能力, 其PRNT50分别为0.01 μ g/mL、0.014 μ g/mL、0.014 μ g/mL和0.023 μ g/mL。结合活性最差的N-5单克隆抗体PRNT50为21.24 μ g/mL。
 | 图3 应用ELISA方法对抗体反应性进行检测Fig.3 Reactivity test of DENV-specific antibodies |
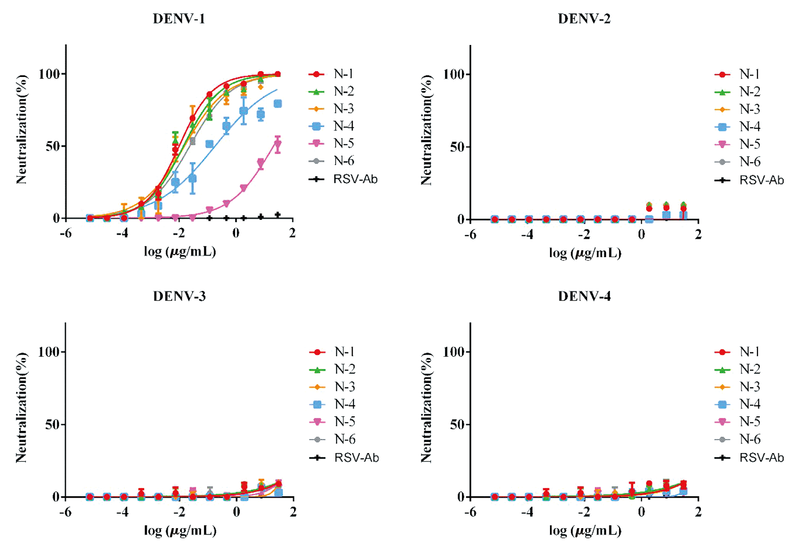 | 图4 测定各抗体对不同亚型DENV的中和活性Fig.4 Neutralization test of DENV-specific antibodies |
配对抗体轻重链可变区基因序列上传至IMGT数据库进行比对并分析。24株抗体轻重链V区基因与人源抗体原始家系一致性均达到90%以上, 说明调取的抗体基因未出现其他物种基因污染。验证为DENV-1 E蛋白特异性的6株单克隆抗体的轻重链基因序列分析结果见表1, 其重链V基因家系为1-69、5-51和4-30, 轻链V基因家系来源为2-14、3-15、1-51、1-39和1-47。与结合活性结果共同分析可以发现, 高结合活性的N-1、N-2、N-3和N-64株单抗均来源于1-69家系。同时这4株单抗均有相似的CDR3氨基酸序列, 可能识别相同的E蛋白抗原表位, 且通过结合该表位能够高效的抑制病毒对细胞的感染。
| 表1 不同抗体的轻重链VDJ基因家系及CDR3氨基酸序列分析 Tab.1 Analysis of VDJ gene family of light and heavy chains with different antibodies, which was accessible from IMGT |
治疗性单克隆抗体的商业开发在20世纪80年代初开始, 目前单克隆抗体药物已经被批准用于治疗多种疾病, 范围从呼吸道合胞病毒感染, 到某些癌症和多发性硬化症, 甚至哮喘和类风湿性关节炎等疾病[21, 22, 23]。在过去25年, 超过30种IgG及其衍生物已被批准用于临床应用[24]。随着研究者对治疗性单克隆抗体研究的不断深入, 其在治疗多种病毒的感染中发挥越来越重要的作用, 尤其在呼吸道合胞病毒的治疗应用中[25, 26]。近几年, 新兴的单个 B 细胞抗体制备技术是一种体外克隆和表达单个抗原特异性 B 细胞抗体基因技术, 这种方法保留了轻重链可变区的天然配对, 具有基因多样性好、效率高、全人源、需要的细胞量少等优势[15, 17, 27]。
本研究主要通过应用抗原特异IgG+记忆B细胞流式筛选平台, 对DENV感染者PBMC中的E蛋白特异IgG+记忆B 细胞进行分选。通过单细胞PCR共得到24对轻重链配对抗体基因, 经过表达后鉴定, 其中6株单克隆抗体为E蛋白特异性的人源单克隆抗体。筛选得到的单克隆抗体与4种亚型的E蛋白结合活性存在明显差异, 与DENV-1 E蛋白的结合活性最强。蚀斑减少中和试验检测结果表明, 筛选得到的单克隆抗体对DENV-1均具中和活性。其中N-1、N-2、N-3和N-6的DENV-1 病毒中和能力最强, 该4株均来源于1-69基因家系, 具有相似的CDR3氨基酸序列, 识别DENV-1 E蛋白上某一表位发挥很强的病毒中和能力。本研究在抗原特异性记忆B细胞中, 成功筛选出具有高中和活性的抗DENV-1人源单克隆抗体。这些抗体不仅能够作为开展DENV研究的重要工具, 而且也可能成为用于开发诊断及临床治疗的重要单克隆抗体。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|