





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
家蝇抗菌肽MAF-1A体外抗甲型流感病毒活性及机制研究
[朱志翠1, 2 , 邓思波1, 2 , 张迎春1 , 吴坤1, 2 , 马晓琳1, 2 , 吴建伟1 , 王涛1, 2  ]
]
]
|
|


The purpose of our research was to explore the effects and potential mechanisms of antimicrobial peptide MAF-1A from Musca domestica on anti-influenza A virus (IAV) activity. In this research, activity of MAF-1A against IAV in vitro were detected with the CPE, MTT assay and quantitative real-time PCR. Cytotoxicity of MAF-1A on MDCK cells was determined by MTT assay. Transmission electron microscope (TEM), hemagglutination inhibition assay and neuraminidase inhibition assay were used to investigate the mechanism for its anti-IAV activities. MAF-1A could significantly inhibit the infection of IAV in vitro, and its median effective concentration (EC50) was (89.8±2.97)μg/mL, and the low cytotoxicity showed on MDCK cells. The results of TEM indicated that MAF-1A had an obvious destructive effect on IAV. The hemagglutination induced by IAV was inhibited by MAF-1A concentration of 1.56μg/mL. MAF-1A had the inhibition on NA (IC50 (134.7±10.31)μg/mL). This study suggests that MAF-1A have remarkably inhibitory effects on IAV, the mechanisms may be associated with the virion damage, combination with hemagglutinin (HA) HA1 subunit and neuraminidase inhibition, which suggest that the anti-IAV of MAF-1A may be through multi-targets.
流行性感冒是一种急性呼吸道传染病, 多由甲型流感病毒(influenza A virus, IAV)引起。由于全球每年感染的病例数量庞大、传播速度快且病毒变异性强, 流感已成为严重威胁公众健康的病毒性传染病之一[1, 2]。目前, 对流感的主要防治措施主要是疫苗接种和化学药物治疗。流感病毒变异性强, 增加了流感疫苗制备的难度, 也使疫苗的有效性大大降低[3, 4, 5, 6, 7]。随着耐药病毒株的不断出现, 现有的抗流感病毒药物已无法满足临床需求[8, 9, 10, 11]。因此, 寻求新型抗流感病毒药物已成为迫切需要解决的问题。
抗菌肽(Antimicrobial peptides, AMPs)是生物体内对细菌、病毒、真菌等微生物和肿瘤细胞具有抑杀活性的小分子多肽。由于具有广谱抗微生物活性、低毒、不容易产生耐药性等特征, AMPs被认为是新型抗生素的理想候选者[12, 13]。MAF-1A(Musca domestica antimicrobial peptide-1A)来源于家蝇, 由26个氨基酸残基构成的小分子抗菌肽[14]。实验研究发现, MAF-1A具有抗流感病毒作用, 具有研发新型抗流感病毒药物的潜力, 但MAF-1A抗流感病毒活性及其作用机制尚未清楚[15]。本文在此基础上, 探讨MAF-1A抗IAV活性和作用机制, 为MAF-1A的应用研究奠定基础。
1.1.1 MAF-1A的化学合成 抗菌肽MAF-1A序列为KKFKETADKLIESAKQQLESLAKEMK, 委托生工生物工程(上海)股份有限公司采用FMOC固相合成法合成, 高效液相色谱(HPLC)纯化、液相色谱-质谱(LC-MS)验证, 多肽合成纯度≥ 98%。
1.1.2 细胞和病毒 狗肾细胞(MDCK细胞)购自中国医学科学院基础医学院研究所; 甲型流感病毒A/PR/8/34 (H1N1) 为本室保存。
1.1.3 主要试剂 胎牛血清购自德国PAN公司; 神经氨酸酶抑制剂筛选试剂盒购自上海碧云天生物技术有限公司; RNAiso Plus试剂盒、逆转录试剂盒(PrimeScriptTM RT Master Mix)、荧光定量PCR试剂盒(SYBR Premix Ex TaqTMⅡ )均购自TaKaRa(大连)有限公司; 四甲基偶氮唑盐(MTT)购自北京博奥拓达科技有限公司; 磷酸奥司他韦购自上海麦克林生化科技有限公司; 其他试剂均为国产分析纯。
1.1.4 引物合成 委托生工生物工程(上海)股份有限公司合成IAV的qRT-PCR检测基因和内参基因引物HA-F:CCTG CTCGAAGACAGCCACAACG, HA-R:GCTCCCTCAGCTCCTCATAGTCG; GAPDH-F:AGGGCAATGCCAGCCC-CAG CG, GAPDH-R:AGGGCAATGCCAGCCCC-AGCG。
1.2.1 MAF-1A抗IAV活性检测
1.2.1.1 病毒的增殖 按文献[16]方法, 将IAV稀释液接种于9日龄鸡胚尿囊腔, 37 ℃培养箱内孵育至48 h, 收获尿囊液, 检测血凝效价。
1.2.1.2 MDCK细胞培养 MDCK细胞用含10%胎牛血清的DMEM培养基在37 ℃ 5% CO2培养箱中培养, 待细胞生长至70%~90%, 取0.5%胰蛋白酶进行消化成单个细胞, 按2× 104个∕ 孔接种于96孔板, 待细胞生长至90%单层细胞时备用。
1.2.1.3 病毒对MDCK细胞半数感染量(TCID50)测定 将上述96孔板生长的MDCK细胞, 去培养液, PBS洗涤3次, 将病毒液以10倍系列稀释, 分别以100 μ L/孔接种于单层MDCK细胞, 2 h后去掉病毒液, PBS缓冲液洗涤3次, 加入等量细胞维持液, 每个稀释度重复6个复孔, 设细胞培养组为阴性对照, 48 h后观察并记录细胞的病变效应; 并参照文献[17]方法, 按Reed-Muench法计算病毒对细胞的半数感染量TCID50。试验共重复3次。
1.2.1.4 MAF-1A对IAV半数有效浓度(EC50)检测 取100× TCID50病毒稀释液50 μ L与等体积MAF-1A溶液混匀, 使MAF-1A终浓度为15.6 μ g/mL、31.2 μ g/mL、62.5 μ g/mL、125 μ g/mL、250 μ g/mL、500 μ g/mL。将混合液于37 ℃预处理30 min后, 接种于单层MDCK细胞, 2 h后去除液体, PBS缓冲液洗涤3次, 以100 μ L/孔加入细胞维持液, 于37 ℃、5% CO2培养箱中培养48 h, 镜下观察细胞CPE效应; MTT法检测MAF-1A抗病毒有效率。同时, 设置细胞培养组和病毒组为对照组, 奥司他韦为药物对照组, MAF-1A对流感病毒抑制率(%)按公式计算, 并参考文献[18], 采用Probit回归法计算半数有效浓度EC50:
有效率=[(药物组OD-病毒组OD)/(阴性对照组OD-病毒组OD)]× 100%
1.2.1.5 qRT-PCR检测MDCK细胞内的病毒载量 取100× TCID50病毒稀释液50 μ L与等体积MAF-1A溶液混匀, 使MAF-1A终浓度为250 μ g/mL、125 μ g/mL、62.5 μ g/mL、31.2 μ g/mL、15.6 μ g/mL。将混合液于37 ℃预处理30 min后, 接种于6孔板中90%单层MDCK细胞, 2 h后去混合液, 用PBS缓冲液洗涤3次, 每孔加入100 μ L细胞维持液, 于37 ℃, 5% CO2培养箱中继续培养24 h后, 根据 TaKaRa公司的 RNAiso Plus 和PrimeScriptTM RT Master Mix试剂盒使用说明书, 提取细胞Total RNA并逆转录为cDNA, 于-20 ℃保存备用。参考文献[19], 使用qRT-PCR 反应试剂盒以管家基因GAPDH为内参, 采用2-Δ Δ Ct 法对HA基因 mRNA 水平进行相对定量。反应条件为:95 ℃ 30 s, 95 ℃ 5 s, 60 ℃ 30 s, 共40个循环。
1.2.2 MAF-1A对 MDCK细胞的毒性检测 将MDCK细胞以2× 104个/孔接种于96孔板, 待其长成90%单层细胞备用; 将MAF-1A溶液进行倍比稀释至10~2 500 μ g/mL, 以上浓度梯度的MAF-1A以每孔100 μ L加入细胞中, 每个浓度设置6个复孔, 并设正常对照组(只加入细胞维持液), 置CO2培养箱中培养至24 h后倒置显微镜下观察 CPE。弃孔内培养基后用 PBS 洗涤 3 遍, 以10 μ L/孔加入5 mg/mL的MTT液, 继续培养4 h后去上清液, 每孔加入DMSO 150 μ L, 振荡摇匀15 min, 在酶标仪490 nm波长下测定各孔吸光度值(OD), 按公式计算 MAF-1A对MDCK细胞的抑制率[18]。
抑制率=[1-(药物处理组OD值-调零孔OD值) /(阴性对照组OD值-调零孔OD值)]× 100%。
1.2.3 MAF-1A抗流感病毒作用机制分析
1.2.3.1 MAF-1A对IAV形态结构的影响 按文献[20]方法, 采用醛化鸡红细胞吸附释放法对含甲型流感病毒A/PR/8/34 (H1N1) 鸡胚尿囊液进行纯化, 并检测纯化后病毒的血凝效价。将纯化的病毒液50 μ L与等体积MAF-1A系列稀释液充分混合, 37 ℃孵育30 min, 用0.1%甲醛溶液于37 ℃灭活48 h, 以2%磷钨酸负染色液进行染色, 经透射电镜观察病毒形态结构。
1.2.3.2 血凝抑制试验 将25 μ L不同浓度的MAF-1A 与等体积4倍血凝效价的病毒液混合, 于室温孵育30 min后, 每孔加入1% cRBC悬液50 μ L, 室温(20 ℃~25 ℃)静置40 min后观察结果, 每个浓度作3个复孔。同时, 设置病毒组和PBS阴性对照组, 试验重复3次。
1.2.3.3 神经氨酸酶抑制试验 按试剂盒说明书操作, 在96孔荧光酶标板内每孔加70 μ L神经氨酸酶检测缓冲液, 然后每孔加入 10 μ L 神经氨酸酶, 再每孔加入10 μ L各浓度的MAF-1A, 振动摇匀 1 min 后于 37 ℃ 孵育2 min; 加入10 μ L 的神经氨酸酶荧光底物, 再振荡 1 min, 37 ℃ 孵育 20 min 进行荧光测定。根据试剂盒方法计算出样品对神经氨酸酶的抑制百分比。
1.2.4 统计学处理 数据以(
经检测发现, 甲型流感病毒A/PR/8/34 (H1N1) 在鸡胚中扩增后, 其血凝效价为1∶ 1 280; 甲型流感病毒A/PR/8/34 (H1N1) 对MDCK细胞的TCID50为1.0× 10-5.2/mL。
MTT结果显示, 当MAF-1A的浓度达2 500 μ g/mL时, 仍对MDCK细胞的生长无明显抑制作用; 镜下可见, 经MAF-1A作用后细胞的形态和结构完整性与对照组相比未见明显差异。MAF-1A 对MDCK细胞表现出较小的细胞毒性。
镜下可见, 甲型流感病毒A/PR/8/34 (H1N1) 可导致 MDCK 细胞变圆, 坏死脱落, 漂浮在培养液表面。但随MAF-1A浓度的增高, 甲型流感病毒A/PR/8/34 (H1N1) 病毒所致的CPE不断减弱, 当浓度达500 μ g/mL时, MDCK细胞完整性好, 形态规则, 正常细胞数量与阴性对照组无明显差异(图1)。经MTT法检测, MAF-1A具有抗甲型流感病毒A/PR/8/34 (H1N1) 活性, MAF-1A对病毒的EC50值为(89.8± 2.97)μ g/mL, 且浓度越大, 对甲型流感病毒A/PR/8/34 (H1N1) 的抑制率越高(图2)。qRT-PCR检测结果显示, 与IAV组比较, 不同剂量(250 μ g/mL、125 μ g/mL、62.5 μ g/mL、31.2 μ g/mL、15.6 μ g/mL)的MAF-1A显著抑制了IAV HA mRNA的表达(t=7.39、6.66、5.78、4.75、5.26, P< 0. 01), 并呈剂量依赖性(图3)。
 | 图1 MAF-1A抑制流感病毒A/PR/8/34 (H1N1) 在MDCK细胞中诱导的CPE(× 400) MAF-1A浓度为A:0 μ g/mL; B:125 μ g/mL; C:500 μ g/mL; D为阴性对照组。Fig.1 MAF-1A inhibited the influenza A/PR/8/34 (H1N1) virus induced CPE in MDCK cells(× 400) |
 | 图2 MAF-1A抗甲型流感病毒A/PR/8/34 (H1N1)活性Fig.2 Inhibition rate of the influenza A/PR/8/34 (H1N1) virus by MAF-1A |
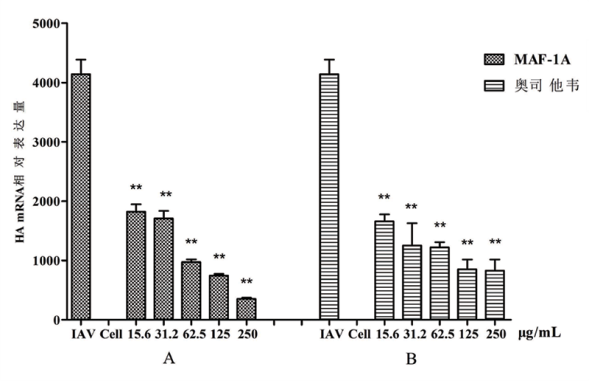 | 图3 MAF-1A处理后甲型流感病毒HA基因mRNA水平的变化 (A)MAF-1A(μ g/mL) (B)奥司他韦 注:* * 与IAV组比较P< 0.01。Fig.3 The mRNA level of influenza A/PR/8/34 (H1N1) virus HA gene after treatment with MAF-1A |
透射电镜检测结果显示, 未经药物处理的病毒对照组病毒颗粒结构完整, 而在MAF-1A的作用下, 病毒的结构出现变化, 随着MAF-1A作用浓度升高, 变化更加明显, 当浓度达500 μ g/mL时, 病毒颗粒被完全破坏(图4)。
 | 图4 不同浓度MAF-1A对甲型流感病毒H1N1形态结构的影响(× 30 000) MAF-1A浓度为A: 0 μ g/mL; B: 125 μ g/mL; C:250 μ g/mL; D:500 μ g/mL。Fig.4 Effects of MAF-1A on morphological structure of virus(× 30 000) |
MAF-1A对流感病毒引起的鸡红细胞凝集具有明显抑制作用, 当浓度为1.56 μ g/mL时即可抑制流感病毒引起的鸡红细胞凝集(图5A)。而MAF-1A与鸡红细胞单独作用时, 不会引起鸡红细胞的凝集(见图5B)。
 | 图5 MAF-1A血凝抑制试验 A:实验组; B:MAF-1A对照组Fig.5 Hemagglutinin inhibition assay for MAF-1A |
结果显示, MAF-1A与阳性对照药物奥司他韦相似, 也具有抑制甲型流感病毒神经酸酶的作用, MAF-1A浓度增高, 抑制效果越明显, 其半数抑制浓度IC50为(134.7± 10.31)μ g/mL(图6)。
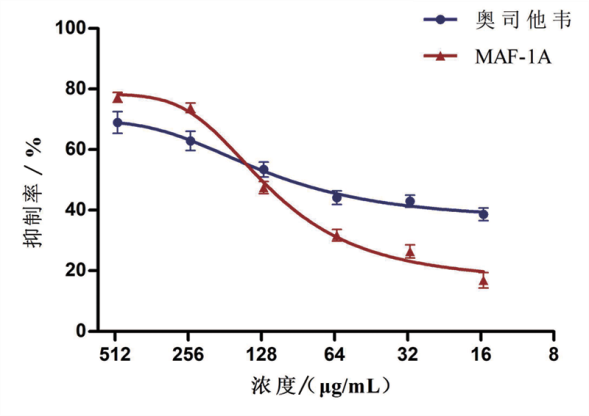 | 图6 MAF-1A对神经氨酸酶活性的影响Fig.6 Neuraminidase inhibition assay for MAF-1A |
目前流感的防治主要依靠药物和疫苗, 但由于耐药病毒株的不断出现以及疫苗作用的局限性, 探寻新型、有效的抗流感药物已成为必然。MAF-1A 是来源于家蝇幼虫体内的一类新型小分子AMPs, 实验研究发现其对多种病毒、真菌具有的抑杀活性, 具有新型肽类药物的开发潜能[15]。本实验研究结果显示, MAF-1A可抑制甲型流感病毒A/PR/8/34 (H1N1) 在MDCK细胞中诱导的CPE, 能有效降低MDCK细胞内的病毒载量; MAF-1A对甲型流感病毒A/PR/8/34 (H1N1) 的EC50值为(89.8± 2.97)μ g/mL, 其对病毒的抑制作用呈剂量相关性, 而MAF-1A对MDCK细胞的毒性小。本实验结果证实, MAF-1A具有较强的抗甲型流感病毒A/PR/8/34 (H1N1) 活性, 且毒性较低, 表明其具有进一步研究的价值。
探明AMPs的抗病毒作用机制对研究AMPs生物学特性、开发新的抗病毒药物非常重要。研究报道, BPI、LL-37等抗菌肽可破坏病毒颗粒的结构, 直接杀灭流感病毒[21, 22, 23]。本实验通过透射电镜观察发现, MAF-1A在低浓度下即可改变流感病毒的电子密度, 随着浓度增加, 对病毒结构破坏越明显, 表明MAF-1A能够直接破坏流感病毒形态结构的完整性, 导致其不能正常复制增殖。
流感病毒的包膜蛋白包括HA和NA。病毒进入靶细胞是由病毒包膜上的 HA介导的。HA 由 HA1 和 HA2 两个亚基构成, 其中 HA1亚基与易感细胞膜表面的唾液酸受体结合, 病毒通过胞吞作用进入细胞内。有文献报道, AMPs可以通过与流感病毒HA相互作用, 阻断其与宿主细胞的吸附或融合过程, 从而有效抑制流感病毒感染[24, 25, 26]。本实验发现, MAF-1A能阻止血凝素 HA1 亚基的血凝作用, 因此认为血凝素HA1亚基是其抗流感病毒的作用位点, MAF-1A可能通过与血凝素HA1亚基结合而阻止流感病毒吸附宿主细胞。
流感病毒由细胞内出芽释放时, 需借助自身的 NA 切断病毒蛋白和宿主细胞表面唾液酸之间的连接, 才能使子代病毒释放至胞外, 感染其他宿主细胞[21, 22, 23, 24, 25]。奥司他韦是NA 抑制剂, 选择性地抑制其活性, 干扰流感病毒从细胞的释放, 从而达到抗流感病毒的作用[27]。实验结果显示, MAF-1A也能抑制NA的活性, 表明对NA的抑制可能也是其抗流感病毒作用机制之一。
综上, MAF-1A不仅能够直接破坏流感病毒的结构, 还可以阻止流感病毒的吸附, 干扰流感病毒的释放, 提示MAF-1A是通过对多靶点作用而发挥抗流感病毒效应, 但其抗流感病毒关键机制及其与靶点的结合模式有待进一步研究。本研究为MAF-1A的深入探讨和开发、利用提供了实验依据。
利益冲突:无
引用本文格式:朱志翠, 邓思波, 张迎春, 等.家蝇抗菌肽MAF-1A体外抗甲型流感病毒活性及机制研究[J].中国人兽共患病学报, 2019, 35(9):791-796. DOI:10.3969/j.issn.1002-2694.2019.00.085
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|